Korcan Ayata, Bazel Üniversitesi’nde (İsviçre), BiyoTıp Bölümünde Araşatırmacıdır.
Solunum yollarında bağışıklığın nasıl çalıştığı ve nasıl çalışmadığını elimden geldiğince basitleştirilmiş şekilde anlatacağım.
Sağda solda duyduğunuz, bağışıklık kısa sürüyor, tekrar ve ağır hasta olunuyor, antikorlar 3-5 ayda kayboluyor gibi fantezilerden bahsetmeyeceğim.
Fantezi diyorum çünkü bilimsel temeli eksik veya bağışıklık bilgisi yetersiz kişilerin yargılarını ancak böyle adlandırabilirim.
Solunum yollarında patojenlere karşı evrimleşmiş üç ana unsur bulunur. Her insan ilk ikisiyle doğar, üçüncüyü ise geliştirir.
Duvar
Doğuştan gelen bağışıklık
Uyarlanabilir bağışıklık
– Duvar dediğimiz, tek katmanlı bir epitel hücre dokusu ve bu yapının üzerini kaplayan mukus tabakasından oluşur.
Epitel hücreler silia dediğimiz tüy benzeri uzantılara sahiptir ve bunları ileri geri hareket ettirerek mukusu yavaş yavaş yukarı iteler ve yutaktan sindirim sistemine yönlendirir. Bu şekilde bir yıl boyunca soluduğumuz her türlü patojen ve ortalama 1,8 kilo toz partikül
akciğerden atılır. Mukus içerisinde doğrudan patojenleri hedefleyen, bağlanıp etkisiz hale getiren proteinler ve antikorlar bulunur. Beslenme ve havanın nemine dikkat edilirse bu koruyucu duvar son derece etkilidir. Birçok patojen bu engeli geçemez. https://annualreviews.org/doi/abs/10.1146/annurev-virology-012420-022445
– Doğuştan gelen bağışıklık:
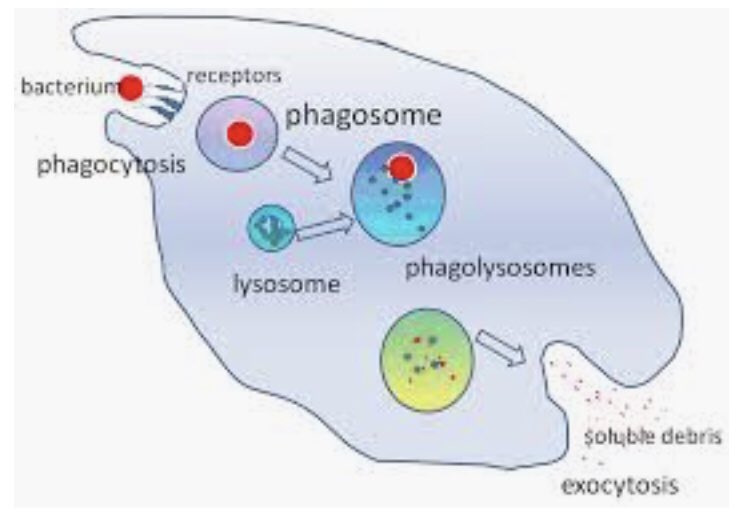
Bu kısım fiziksel bariyer etrafında dolaşan fagosit dediğimiz, “patojen yiyen” hücreleri içeriyor. Makrofaj, monosit, nötrofil gibi. Bunlar antikora ihtiyaç olmaksızın, bakteri, mantar, virüs ve parazitleri tanıyabilir, saldırabilir ve yutabilirler.
– Uyarlanabilir bağışıklık:
En gelişmiş ve kapsamlı bağışıklık bu kategori içindedir. Her bireyin kendi bağışıklığı vardır ve başka kimseye benzemez.
İçinde üç ana grup hücre vardır:
B ve plazma hücreleri
T hücreleri
Antijen sunan hücreler (dendritik hücre ve makrofajlar).
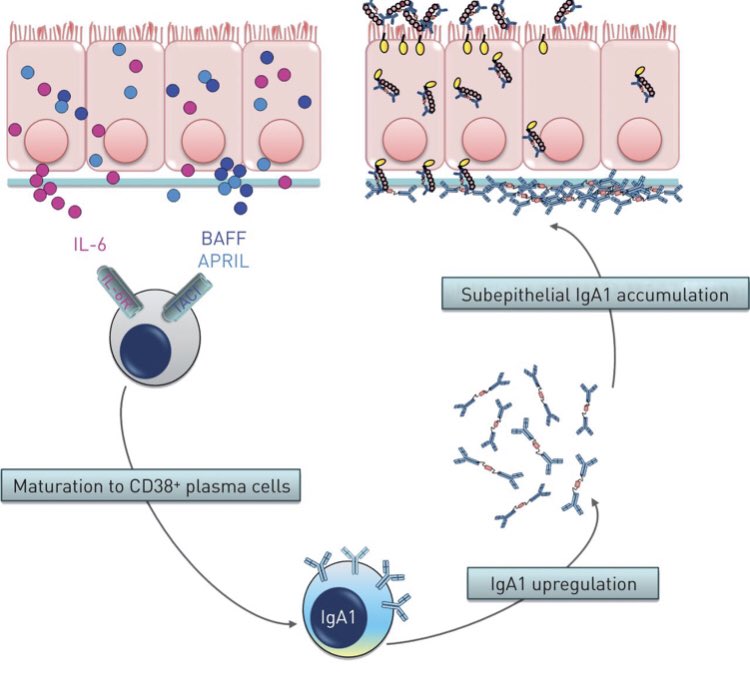
B ve plazma hücreleri solunum yolları mukozası içerisine antikorlar salarlar. Mukozaya özel bu antikorlar IgA tipindedir. Birbirine bağlı iki antikordan yapı 4 bağlanma ucuna sahiptir. Patojenleri çapraz bağlayıp, etkisiz hale getirebilir. Enflamasyon tetiklemez.
IgA tipi antikorlar kanda dolaşan IgG ve IgM antikorlardan yapısal olarak farklı oldukları için daha dayanıklı ve uzun ömürlüdürler. Bu antikorlara kanda az rastlanır. Çünkü bunların görevi mukozada bulunmak ve patojenleri burada tutmaktır.
Patojene özel B ve plazma hücreleri hafızaya sahiptir, yani tamamen aynı veya benzer patojen ile tekrar karşılaşıldığında, hızlı bir şekilde antikor salgılayıp enfeksiyonun yayılmasını engellerler. Bunlar uzun ömürlüdür ve saklandıkları özel cepler vardır. Kanda ender bulunur.
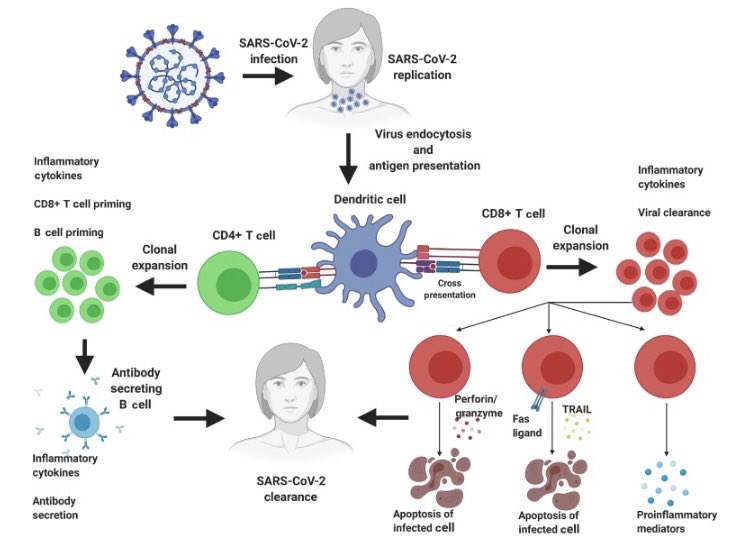
T hücreleri 2 ana kategoriye ayrılır: CD4 ve CD8
CD4 ler “yardımcı hücre” olarak da adlandırılırlar, B hücrelerinin patojenlere karşı hassasiyet kazanmasına ve hedefe özel antikor üretmelerine yardim eder. Gerektiğinde yeni B hücrelerinin gelişimini yönlendirirler.
CD8 ler ise öldürücü lenfosit olarak adlandırılırlar. Virüsle enfekte olmuş veya kanserleşmiş hücreleri tespit edip öldürmek için uzmanlaşmışlardır. Bu sayede virüslerin hücre içinde çoğalması engellenir. Bu hücreler tüm vücut hücrelerine virüs kontrolü yapabilirler.
Antijen sunan hücrelerin görevi, mukozada veya vücut içinde tespit ettikleri yabancı yapıları lenf bezlerine götürüp CD4 yardımcı T hücrelere tanıtmaktır. Bunu bir tür istihbarat servisi gibi düşünebilirsiniz. Sistem birbirine bağlı bir zincir şeklinde çalışır.
Tek başına mukus ve akış kontrolü bile çok önemli bir bileşendir. Bu noktaya kadar, kişinin vücuduna virüs girememiştir, ama süprüntü testinde PCR pozitif çıkar. İçeriye giremeyip kalan virüsler parçalanır ve antijen sunan hücreler tarafından arşivlenir.
Yukarıdaki bileşenlerin herhangi birinde önemli bir bozulma, enfeksiyonlara karşı duyarlılığa yol açar.
Bir bileşen başarısız olunca, işareti kandaki antikorlardır. Tam da bu yüzden, hastalık şiddeti ve virüsün sistemik yayılması, kandaki antikor seviyelerinin göstergesidir.
Birincil savunmalarla daha bir enfeksiyon başlamadan virüsü atlatmış insanlar daha düşük ve daha hızlı azalan sistemik antikor seviyesine sahip olacaktır, bu tamamen normal.
Fakat bu o kişileri kesinlikle korumasız bırakmaz. Artık solunum yolu mukozasinda bağışıklığın kendi hafızası oluşmuştur. Bu hafıza sadece B ve plazma hücreleri için değil, aynı zamanda CD4 ve CD8 T hücre tipleri için de mevcuttur.
Evet, bu hafızalı hücreler onlarca yıl solunum yolu duvarlarında oturup bekleyebiliyorlar, hangi düşmana saldıracaklarını biliyorlar. Beklediklerine oldukça benzer bir patojen bile geldiğinde harekete geçecekler. Bu kalıcı bir bağışıklık hafızası türüdür. Düşman unutulmuyor.
Kandaki antikorlar tamamen yok olsa bile hafızali B hücreleri varlığını sürdürür ve gerektiğinde yeniden antikor üretebilir.
Yeni koronavirüs hakkındaki en önemli sorulardan biri, enfekte olan kişilerin yeniden enfeksiyona karşı bağışık olup olmadığı ve eğer öyleyse bunun ne kadar sürdüğüdür. Yüksek sayıda denekle yapılan çalışmalar bu sürenin en az 6-7 ay olduğunu gösteriyor.
En son yayınlanan ve 30.000 den fazla katılımcı ile yapılan Newyork çalışması da hastalığı geçirenlerin yüksek miktarda bloke edici antikor ürettiğini gösterdi.
Yukarıdakileri okuduysanız, artık kandaki antikorların yalnızca sistemik hastalık durumuna karşı koruyucu olduğunu, üst solunum yolu semptomlarını ve hatta hafif bir zatürreyi bile önleyemediklerini biliyorsunuz; tabi bulaşıcılığı engellemelerinin mümkün olmadığını da.
“Genel olarak, bu bulgular enfeksiyondan altı ay sonra SARS-CoV-2’ye karşı güçlü bir hücresel (T hücresi) bağışıklık tepkisine işaret etmektedir. Bu bulgular, yalnızca SARS-CoV-2’ye karşı bağışıklığın nasıl çalıştığına dair anlayışımızı beslemekle kalmayacak, aynı zamanda… gelecekteki aşı stratejilerine de yardımcı olacaktır. Bu hücresel bağışıklık tepkisinin uzun vadede devam edip etmediği ve yeniden enfeksiyon olasılığına nasıl karşılık vereceğimin araştırılmaya ihtiyacı vardır”
Bu noktada bağışıklık sistemi ve bulaşıcı hastalık uzmanlarının bile bu mekanizmaları görmezden gelip fantazi kıvamında açıklamalar yapmasını hayretle izliyorum. Bağışıklık inkarcılarına kulak asmayın!
En sona da bir video ekleyelim, bağışıklık sistemini basitçe anlatıyor. İngilizce ama konu çok güzel işlenmiş.
SARS-CoV-2 spike proteininin yapısal ve fonksiyonel özellikleri: COVID-19 için potansiyel antivirüs ilaç geliştirme
COVID-19, şu anda dünya çapında hala yayılım gösteren ve yeni ortaya çıkan bulaşıcı bir hastalıktır. Hastalık, yeni bir koronavirüs olan şiddetli akut solunum sendromu koronavirüs 2’den (SARS-CoV-2) kaynaklanmaktadır. SARS-CoV-2’nin Başak (Spike, S) proteini, reseptör tanıma ve hücre zarı ile füzyon (kaynaşma) sürecinde önemli rol oynar. Spike proteini, S1 ve S2 olmak üzere iki alt birimden oluşur. S1 alt birimi, konakçı “anjiyotensin dönüştürücü enzim-2 reseptörünü” (ACE2) tanıyan ve ona bağlanan bir reseptör bağlanma alanı bölgesi, RBD) içerir. S2 alt birimi, iki heptad (7 amino asitlik) tekrar bölgesi yoluyla altı sarmallı bir demet oluşturarak virüsün hücre membran (zar) füzyonuna aracılık eder.
COVID-19, Aralık 2019’da meydana gelen yeni bir koronavirüsten kaynaklandı. Kısa sürede (5 ay içinde) dünya çapında yayıldı ve küresel bir pandemiye dönüştü. COVID-19’un daha sonra şiddetli akut solunum sendromu koronavirüs 2 (SARS-CoV-2) olarak adlandırılan bir koronavirüsün neden olduğu kısa sürede keşfedildi. Virüs, β koronavirüs ailesine aittir. İnsanları enfekte ettiği bilinen yedinci koronavirüstür; Bu koronavirüslerden dördü (229E, NL63, OC43 ve HKU1) sadece hafif soğuk algınlığı semptomlarına neden olurken, diğer üçü (SARS-CoV, MERS-CoV ve SARS-CoV-2) ciddi semptomlara sebep olmakta ve hatta sırasıyla% 10,% 37 ve% 5 oranları ile ölümlere neden olabilmektedir.
SARS-CoV-2, tek sarmallı, pozitif (+) anlamlı (sens) RNA içeren zarflı bir virüstür. RNA tabanlı metagenomik yeni nesil dizileme yaklaşımı ile, tüm genomunun 29.881 bp uzunluğunda olduğu gösterildi (GenBank no. MN908947). Genom 9860 amino asitii kodlar. Gen fragmanları, yapısal ve yapısalolmayanproteinleri ifade eder. S, E, M ve N genleri yapısal proteinleri kodlarken, 3-kimotripsin benzeri proteaz, papain benzeri proteaz ve RNA’ya bağımlı RNA polimeraz (RdRp) gibi yapısal olmayan proteinler ORF bölgesi tarafından kodlanır.
SARS-CoV-2’nin yüzeyini çok sayıda glikozillenmiş (şeker molekülleri ile kaplı) Spike proteini (S)proteini kaplar. Bu proteinler, viral hücre girişine aracılık ederek, konakçı hücre reseptörü anjiyotensin dönüştürücü enzim 2’ye (ACE2) bağlanır.
S proteini reseptöre bağlandığında, TM proteaz serin 2 (TMPRSS2), S proteinini aktive ederek virüsün hücreye girişini destekler. TMPRSS2 konak hücre membranında bulunan TMPRSS2a tip 2 bir TM serin proteazdır. Virüs hücreye girdiğinde, viral RNA salınır ve poliproteinler ifade edilir. Viral RNA genomunun replikasyonu ve transkripsiyonu, protein işlenmesi, replikaz-transkriptaz kompleksinin birleşmesi yoluyla gerçekleşir. Viral RNA kopyalanır ve yapısal proteinler, konakçı hücrede sentezlenir, birleştirilir ve paketlenir, ardından viral partiküller serbest bırakılır (Şekil 1d).
Şekil: SARS-CoV-2 S proteininin şeması. a) S proteininin şematik yapısı. b) S proteini, reseptör ACE2’ye bağlanır. c) S proteininin aracılık ettiği bağlanma ve virüs-hücre füzyon süreci. d) SARS-CoV-2’nin konukçu hücrelerdeki yaşam döngüsü.
Bu proteinler, viral yaşam döngüsü için kritiktir ve ilaç tedavileri için potansiyel hedeflerdir. Örneğin, ACE2 bazlı peptid, 3CLpro inhibitörü (3CLpro-1) ve yeni bir vinilsülfon proteaz inhibitörünün SARS-CoV-2’ye karşı etkili olduğu deneysel olarak gösterilmiştir. SARS-CoV-2’nın S proteini, tüm insan koronavirüsleri (HCoV’ler) arasında yüksek oranda korunur ve reseptör tanıma, viral bağlanma ve konakçı hücrelere girişte rol oynar. Vazgeçilmez fonksiyonları nedeniyle COVID-19 aşısı ve terapötik araştırmalar için en önemli hedeflerden birini temsil etmektedir.
S proteininin yapısı
180–200 kDa boyutundaki S proteini, hücre dışı bir N-ucu, viral membrana sabitlenmiş bir transmembran (TM) bölgesi ve kısa bir hücre içi C-ucu segmentinden oluşur. S proteini normalde yarı kararlı, prefüzyon (birleşme öncesi) yapısında bulunur. Virüs, konakçı hücre ile etkileşime girdiğinde, S proteininde kapsamlı yapısal bir yeniden düzenlenme meydana gelir ve virüsün konakçı hücre membranı ile kaynaşmasına izin verir. Proteinin sivri çıkıntıları, kamufle etmek ve böylece bağışıklık tepkisinden korunmak için polisakkarit molekülleri ile kaplanır ve böylece hücreye giriş sırasında virüs konakçı bağışıklık sisteminin denetiminden kaçınır.
SARS-CoV-2 S’nin toplam uzunluğu 1273 amino asittir. N-ucunda bulunan bir sinyal peptidinden (amino asitler 1-13), S1 alt biriminden (amino asitler 14-685) ve S2 alt biriminden (amino asitler 686–1273) oluşur; son iki bölge sırasıyla reseptör bağlanmasından ve membran füzyonundan sorumludur. S1 alt biriminde, bir N-terminal alanı (14–305 amino asitler) ve bir reseptör bağlanma alanı (RBD, 319–541 amino asitler) vardır; füzyon peptidi (FP) (788–806 amino asitler), heptapeptid tekrar dizisi 1 (HR1) (912–984 amino asitler), HR2 (1163–1213 amino asitler), TM alanı (1213–1237 amino asitler) ve sitoplazma alanı (1237– 1273 amino asitler), S2 alt birimini içerir (Şekil 2a). S protein trimerleri (üç eşdeğerdeki birimi), görsel olarak viral partikülü çevreleyen karakteristik soğanlı, taç benzeri bir hale oluşturur (Şekil 1a). Koronavirüs S protein monomerlerinin yapısına bağlı olarak, S1 ve S2 alt birimleri soğan başı ve sap bölgesini oluşturur. SARS-CoV-2 trimerik S proteininin yapısı, atomik seviyede kriyo-elektron mikroskobu (soğutmalı elektron mikroskobu) ile belirlenmiş ve S reseptör bağlanma bölgesi (RBD) açık ve kapalı durumdaki şekillerini ve işlevlerini ortaya çıkarmıştır (Şekil 2b, c).
Şekil: SARS-CoV-2 S proteininin yapısı. SARS-CoV-2 spike proteininin yapısal ve fonksiyonel özellikleri ve COVID-19 için potansiyel antivirüs ilaç geliştirme. a. SARS-CoV-2 proteini başlığının şematik gösterimi. b – c. S proteini RBD kapalı ve açık durumu. d. S proteini, S1 alt biriminde açılmış RBD ile ACE2’ye bağlanır. e. S2 alt biriminin HR1 ve HR2 tarafından oluşturulan altı sarmallı yapı.
Doğal durumda, S proteini inaktif bir öncü olarak bulunur. Viral enfeksiyon sırasında hedef hücre proteazları, S proteinini S1 ve S2 alt birimlerine bölerek etkinleştirir. Bu bölünme, hedef hücrelere viral girişten sonra membran füzyon bölgesini aktive etmek için gereklidir. Diğer koronavirüslere benzer şekilde, SARS-CoV-2’nin S proteini, hücresel proteazlar tarafından S1 ve S2 alt birimlerine serin proteaz TMPRSS2 ile bölünür. SARS-CoV’nin bölünme bölgesi bilinmesine rağmen, SARS-CoV-2 S’ninki henüz rapor edilmemiştir.
S1 alt biriminin yapısı
Virüs partiküllerinin konakçı hücrenin yüzeyindeki hücre reseptörlerine bağlanması, virüs enfeksiyonunu başlatır; bu nedenle reseptör tanıma, viral girişin önemli bir belirleyicisi ve bir ilaç tasarım hedefidir.
S1 alt biriminde yer alan RBD, hücre reseptörü ACE2’de aminopeptidaz N bölgesine bağlanır. S1 bölgesi, N-terminal bölgesi (NTD) ve C-terminal bölgesi (CTD)’yi içerir ve bağlanma arayüzündeki atomik ayrıntılar, SARS-CoV-2-CTD’deki anahtar amino asitlerin ikamelerini gösterir. Ek olarak, SARS-CoV-2 S CTD bağlanma arayüzü, SARS-RBD’den ACE2 reseptörü ile doğrudan etkileşime giren daha fazla amino asite sahiptir (21’e karşı 17) ve SARS-CoV-2 S CTD ile daha geniş bir yüzey alanı gömülüdür. Anahtar amino asitlerin mutasyonları, ACE2 ile etkileşimin güçlendirilmesinde önemli bir rol oynar. SARS RBD’de I472 yerine SARS-CoV-2’deki F486, ACE2 Y83 ile güçlü aromatik-aromatik etkileşimler oluşturur ve SARS-CoV-2-CTD’de E484, SARS RBD’de P470 yerine K31 ile iyonik etkileşimler oluşturur. Bu durum, SARS-CoV’nin RBD’sine göre reseptör bağlanması için daha yüksek afiniteye yol açar (Şekil 2d).
RBD bölgesi, nötralize antikorlar (nAb’ler) için kritik bir hedeftir ve SARS-CoV-2’daki bu bölge SARS-CoV RBD ~% 73-% 76 benzerdir. CoV RBD’de ACE2 ile temas eden dokuz amino asit tamamen korunurken ve dördü kısmen korunur. SARS-CoV ve SARS-CoV-2’nin RBM’sinin (reseptör bağlama motifi, RBD’nin bir kısmının ACE2 ile doğrudan temas kurması) analizi, SARS-CoV S proteinindeki ACE2 bağlanması için gerekli olan çoğu amino asitin SARS’ta korunduğunu ortaya çıkarmıştır. Bununla birlikte, bazı çalışmalar, fare monoklonal antikorlarının (mAb’ler) ve SARS-RBD’ye karşı poliklonal antikorların SARS-CoV-2’nin S proteini ile etkileşime giremediğini ve SARS-CoV ve SARS-CoV-2 arasındaki antijenisitede farklılıkları ortaya çıkardığını göstermiştir. Benzer şekilde, SARS-CoV RBD’ye özgü bir antikor, SL-CoV-SHC01’in S proteininin aracılık ettiği enfeksiyonu bloke edemedi, bu da S1 RBD’nin, geniş spektrumlu ve oldukça değişken karakteristiği nedeniyle ideal bir ilaç hedefi olmayabileceğini düşündürmektedir.
S2 alt biriminin yapısı
Ardışık olarak bir FP, HR1, HR2, TM bölgesi ve sitoplazmik alan füzyonundan (CT) oluşan S2 alt birimi, viral füzyon ve girişten sorumludur.
FP, viral ailenin 15-20 korunmuş amino asitlerden oluşan kısa bir bölümüdür ve esas olarak glisin (G) veya alanin (A) gibi hidrofobik amino asitleri içerir. FP peptidleri, S proteini ön saç tokası yapısını benimsediğinde hedef zara tutunur. Önceki araştırmalar, FP’nin, konakçı hücre membranının çift lipit katmanlarını bozarak membran füzyonuna aracılık etmede önemli bir rol oynadığını göstermiştir.
HR1 ve HR2, tekrarlayan bir heptapeptidden (7 amino asitlik bir peptid) oluşur: HPPHCPC, burada H hidrofobik ve hacimli bir amino asiti simgeler, P polar veya hidrofilik bir amino asiti ve C başka bir yüklü amino asiti simgeler. HR1 ve HR2, S2 alt biriminin viral füzyon ve hücreye giriş fonksiyonu için gerekli olan altı sarmal demeti (6-HB) (Şekil 2e) oluşturur. HR1, bir hidrofobik FP’nin C-ucunda bulunur ve HR2, TM bölgesinin N-ucunda bulunur. Aşağı akış TM alanı, S proteinini viral membrana tutturur ve S2 alt birimi bir CT kuyruğunda son bulur.
RBD, ACE2’ye bağlanır ve S2, FP’yi hedef hücre zarına sokarak konformasyonu değiştirir. Bu, HR1 alanının ön saç pimli sarmal bobinini açığa iter ve 6-HB oluşturmak için HR2 alanı ile HR1 trimer arasındaki etkileşimi tetikleyerek viral zarf ve hücre zarını viral füzyon ve giriş için yakın konuma getirir. HR1, yüzeyde HR2’ye bağlanan yüksek oranda korunmuş üç hidrofobik oluğun açıkta olduğu bir homotrimerik düzenek oluşturur. HR2 alanı, HR1 alanı ile etkileşim için hem sert bir sarmal hem de esnek bir döngü oluşturur. CoV’lerin postfüzyon firkete konformasyonunda, “füzyon çekirdek bölgesi” (sırasıyla HR1core ve HR2core bölgeleri) olarak adlandırılan, sarmal bölge içindeki HR1 ve HR2 alanları arasında birçok güçlü etkileşim vardır.
Heptad tekrarını (HR) hedeflemek, terapötik ilaç calışmalaında en büyük ilgiyi çekmiştir. S proteini, spesifik ilaçların geliştirilmesi için önemli bir hedef protein iken, S1 RBD alanı oldukça değişken bir bölgenin parçasıdır ve geniş spektrumlu antiviral inhibitör gelişimi için ideal bir hedef bölge değildir. Aksine, S2 alt biriminin HR bölgesi, HCoV enfeksiyonlarında önemli bir rol oynar ve HR1 ve HR2 arasındaki etkileşim modu gibi HCoV’ler arasında korunmuştur. Viral proteinlerin korunmuş bölgelerinden türetilen peptitlerin antiviral etkinliği önemli bir çalışma alanıdır. Zarflı virüslerin sınıf I viral füzyon proteinlerinin HR2 bölgesinden türetilen peptitler, viral HR1’e rekabetçi bir şekilde bağlanır ve viral enfeksiyonu etkili bir şekilde inhibe eder. Bu nedenle HR1, SARS-CoV-2 enfeksiyonuna karşı füzyon inhibitörlerinin geliştirilmesi için umut verici bir hedeftir.
S proteininin fonksiyonları
Virüs yüzeyindeki S proteini, enfeksiyonla ilgili önemli bir faktördür. Bu protein, viral girişten sorumlu trimerik sınıf I TM bir glikoproteindir ve her türlü HCoV’de olduğu gibi, HIV (HIV glikoprotein 160, Env), influenza virüsü (influenza hemaglutinin, HA), paramiksovirü (paramiksovirüs F) ve Ebola (Ebola virüsü glikoproteini) gibi diğer virüslerde de mevcuttur. Diğer koronavirüslere benzer şekilde, SARS-CoV-2’nin S proteini, viral enfeksiyon sırasında reseptör tanıma, hücre bağlanması ve füzyona aracılık eder.
Viral zarfın yüzeyinde bulunan S proteininin trimeri, S proteininin reseptöre bağlandığı temel birimdir. S1 bölgesi, esas olarak virüsün reseptöre bağlanmasından sorumlu olan RBD’yi içerirken, S2 alanı esas olarak, virüs füzyonu ile yakından ilgili olan HR1 ve HR2 dahil olmak üzere HR bölgesi içerir.
Reseptör bağlama
Yukarıda bahsedildiği gibi SARS-CoV-2’nin S proteini, ACE2 reseptörü vasıtası ile konakçı hücreye bağlanır. ACE2 bir enzim reseptör olup, anjiyotensin I’i anjiyotensin 1-9’a dönüştüren bir ACE homologudur. ACE2 esas olarak akciğer, bağırsak, kalp ve böbrekte ve alveolar epitel tip II hücrelerde ifade edilir. ACE2 aynı zamanda SARS-CoV için bilinen bir reseptördür. SARS-CoV’nin S proteininin S1 alt birimi, ACE2’ye bağlanarak düşük pH altında viral füzyon aktivitesini tetikleyen endozom oluşumunu teşvik eder (Şekil 1a, b).
S proteini ve ACE2 arasındaki etkileşim, amfibiler (krbağalar), kuşlar ve memeliler gibi farklı türlerde korunmuş bir birincil yapıya sahip olması nedeniyle, SARS-CoV-2’nin ara konakçılarını tanımlamak için kullanılabilir.
S proteini, S1 alt biriminin RBD bölgesi yoluyla ACE2’ye bağlanarak, bir trimer formunda konakçı hücrelere viral bağlanmaya aracılık eder. SARS-CoV-2’nin S proteini, 14.7 nM’lik bir ayrışma sabiti (Kd) ile insan ACE2’ye bağlanır, ancak SARS-CoV’nin S’i 325.8 nM’dir. Bu durum, SARS-CoV-2 S’nin ACE2’ye SARS-CoV’den daha duyarlı olduğunu gösterir. SARS-CoV-2 proteinlerinin tanımlanması yoluyla, araştırmacılar SARS-CoV-2 ve SARS-CoV arasında S’de ~% 24 fark bulurken, RBD’de bu farki ~% 23’tür.
Viral füzyon
Viral füzyon, viral genomun konakçı hücreye (yani hücrelerimize) salınmasıyla sonuçlanan viral membran ve konakçı hücre membranının füzyonunu ifade eder. SARS-CoV-2’nin S1 ve S2 alt birimlerinin bölünmesi, füzyonun temelidir. S proteini, konakçı proteazlar tarafından S1 alt birimi ve S2 alt birimi olmak üzere iki parçaya bölünür ve alt birimler, viral füzyon oluşana kadar kovalent olmayan bir formda bulunur. Araştırmacılar, spesifik furin bölünme bölgesinin SARS-CoV-2’nin bölünme bölgesinde bulunduğunu, ancak diğer SARS benzeri CoV’lerde bulunmadığını bulmuşlardır. SARS-CoV-2 veya SARS benzeri CoV’lerde bölünme bölgesinin mutasyonu, SARS-CoV-2’nin S proteininin parçalanmamış bir durumda var olduğunu ancak diğerlerinin esas olarak bölünmüş durumda olduğunu ortaya çıkarmıştır. SARS-CoV-2’nin S proteini, furin benzeri proteazlar tarafından parçalanma olasılığını artıran ve böylece enfektivitesini artıran birden fazla furin parçalama bölgesine sahiptir. Furin benzeri bölünme bölgesi, yüksek derecede patojenik influenza virüsünde de mevcuttur ve 1997’de Hong Kong’daki kuş gribi salgınında gözlemlendiği gibi patojenisitesiyle ilgilidir. Ek olarak, TMPRSS2 gibi konak hücre proteazları, S protein hazırlığı için gereklidir ve SARS-CoV ve influenza A virüsünün girişinde aktive oldukları gösterilmiştir. Viral S proteinini parçaladığı kanıtlanmış bir başka konak hücre proteazı tripsin’dir. Özet olarak, SARS-CoV-2’nin S proteini, SARS-CoV’ninkine benzerdir ve konakçı hücre proteazları, hem SARS-CoV-2 hem de SARS-CoV’nin S protein bölünmesini teşvik etmek için gereklidir. SARS-CoV-2’nin S proteininde belirli bir furin bölünme bölgesinin varlığı, SARS-CoV-2’nin SARS-CoV’den daha bulaşıcı olmasının bir nedeni olabilir.
6-HB oluşumu viral füzyon için gereklidir. SARS-CoV-2’nin N-ucundaki FP ve S2 üzerindeki iki HR bölgesi viral füzyon için gereklidir. S proteininin bölünmesinden sonra, SARS-CoV-2’nin FP’si açığa çıkar ve viral füzyonu tetikler. Bazı özel ligandların etkisi altında, füzyon proteini konformasyonel (yapısal) bir değişikliğe uğrar ve ardından konakçı hücre zarına eklenir (Şekil 1c). Örneğin, influenza A virüsü için ligand H+ iken, HIV için ligand CCR5 veya CXCR4 gibi bir koreseptördür. Viral membran ile konakçı hücre membranı arasındaki mesafe kısalır ve S proteininin HR1 bölgesi, konakçı hücre membranına çok yakınken, HR2 bölgesi viral membran tarafına daha yakındır. Daha sonra, HR2 tekrar HR1’e katlanır, iki HR bölgesi, füzyon çekirdeğinin antiparaleli altı sarmallı bir yapı oluşturur. Viral membran, konakçı hücre membranına doğru çekilir ve ona sıkıca bağlanır ve iki membran birleşir.
S proteinini hedefleyen potansiyel ilaçlar
Viral enfeksiyonda S proteininin temel rolü, aşı gelişimi, antikor bloke edici tedavi ve küçük molekül inhibitörleri için potansiyel bir hedef olduğunu gösterir. SARS-CoV ve MERS-CoV ile benzerlik göz önüne alındığında, SARS-CoV-2’da S’yi hedefleyen potansiyel nAb’ler (nötralize edici antikorlar) ve inhibitörler aşağıda özetlenmiştir (Şekil 3).
Şekil: SARS-CoV-2 S proteinini hedefleyen potansiyel ilaçlar. a. S proteininin çeşitli epitoplarını hedefleyen potansiyel monoklonal antikorlar (mAb’ler). b. Mevcut SARS-CoV-2 inhibitörlerinin özeti.
SARS-CoV-2’nın S proteinine dayalı antikorlar
S proteini, SARS-CoV-2’nin tüm yapısal proteinlerindeki ana antijen bileşenidir. SAS-CoV-2’nin diğer fonksiyonel proteinlerinden farklı olarak, konakçı immün tepkisinin indüklenmesinden sorumludur ve S proteinini hedefleyen nAb’ler, viral enfeksiyona karşı koruyucu immüniteyi indükleyebilir. SARS-CoV ve MERS-CoV’ye benzer şekilde, SARS-CoV-2’nin nAb’leri üzerine yapılan araştırmalar, esas olarak Ab’leri (monoklonal antikorlar), antijen bağlayıcı fragmanları, tek zincirli değişken bölge fragmanlarını ve S1 RBD, S1’i hedefleyen tek alanlı antikorları (Nbs) içerir ve S2 aracılı füzyonu önlemek için S1 RBD, S1-NTD veya S2 bölgelerini hedeflerler. Şu an, adenovirüs bazlı vektörler ve saflaştırılmış inaktive edilmiş virüs dahil olmak üzere birçok SARS-CoV-2 aşı tipi geliştirme aşamasındadır.
SARS-CoV-2 ve SARS-CoV S proteinleri arasındaki dizi ve çarpıcı yapısal benzerlik, COVID-19’u SARS-CoV S proteinini hedefleyen antikorlarla tedavi etme olasılığını vurgulamaktadır. SARS-CoV-2 RBD ile karşılaştırıldığında SARS-CoV-2, C-terminal bölgesi (SARS-CoV-2-CTD) aracılığıyla hACE2 ile etkileşime girerek reseptör bağlanması için daha yüksek afinite gösterir. RBD, oldukça güçlü nAb yanıtları indükleyebilir ve SARS-CoV-2’ye karşı etkili ve güvenli bir alt birim aşı olarak geliştirme potansiyeline sahiptir. Bağışık farelerden elde edilen SARS-CoV S poliklonal antikorları, SARS-CoV S-MLV’nin (murin lösemi virüsü) istilasını tamamen inhibe ederken, SARS-CoV-2 S-MLV’nin istila oranı ~% 10’a düşürülmüştür. Poliklonal anti-SARS S1 antikoru T62, SARS-CoV S girişini inhibe eder ancak SARS-CoV-2 S psödovirüs partiküllerinin girişini engellemez. Tutarlı bir şekilde, son çalışmalar benzer sonuçlar bildirmiş ve SARS RBD’ye yönelik üç mAb, S230, m396 ve 80R’nin SARS-CoV-2 RBD’ye bağlanamadığını göstermiştir.
Öte yandan, birkaç mAb, SARS-CoV-2’yi etkisiz hale getirmede umut verici sonuçlar göstermiştir. SARS-CoV’ye özgü bir insan mAb’si olan CR3022, SARS-CoV-2 (6,3 nM’lik KD, OctetRED96’da BLI ile ölçülmüştür) ile güçlü bir şekilde bağlanır ve CR3022’nin tek başına veya kombinasyon halinde terapötik olarak geliştirilme potansiyeline sahip olduğunu gösterir. İnsan Ig değişken ağır ve hafif zincirleri ifade eden immünize edilmiş transgenik farelerden hazırlanan S1’i hedefleyen bir mAb’nin, RBD-hACE2 etkileşiminin blokajından bağımsız, bilinmeyen bir mekanizma yoluyla hem SARS-CoV-2 hem de SARS-CoV enfeksiyonlarını nötralize ettiği yakın zamanda gösterilmiştir. Son zamanlarda, birçok bloke edici insan mAb’u (311mab-31B5, 311mab-32D4, 47D11, n3130, n3088, S309, P2C-1F11, P2B-2F6, B38, H4), iyileşmiş COVID-19 hastalarının hafıza B hücrelerinden başarıyla klonlanmıştır. Bu mAb’ler, enfeksiyonu etkili bir şekilde nötralize etmek için özellikle SARS-CoV-2 S’ye bağlanır. Ek olarak, rehabilitasyon sırasında SARS hastalarından veya SARS-CoV S1 ile özel olarak aşılanmış hayvanlardan alınan serumlar, SARS-CoV-2’yi çapraz nötralize edebilir ve S proteini aracılı SARS-CoV-2 girişini azaltabilir (Şekil 3).
Füzyon inhibitörleri
SARS-CoV-2’nin S proteininin stabilitesi SARS-CoV S’den daha düşüktür. Sarbecovirus alt cinsinin çoklu S sekanslarının haritalanması, S2 füzyon bölgesinin S1 alt biriminden daha korunmuş oluğunu ve S1 alt biriminin viral yüzeyde daha fazla açığa çıktığını göstermiştir. SARS-CoV S2 alt birimi, virüs-hücre füzyonuna aracılık etmede ve bunun, HR1 ve HR2’nin 6-HB oluşturmak üzere etkileşime girdiği, böylece virüsün hücre zarına bağlanmasını ve bununla kaynaşmasını sağlayan konakçı hücrelere entegrasyonunda önemli bir rol oynar.
Sıra hizalaması, SARS-CoV-2 HR2’nin SARS-CoV HR2 ile aynı diziye sahip olduğunu gösterir. Bu nedenle, SARS-CoV-2 HR2P (1168-1203 amino asitleri), SARS-CoV-2 füzyonunu ve bir hedef hücreye girişini engellemek için tasarlanmıştır. Şaşırtıcı bir şekilde, HR2P, sırasıyla 0.18 ve 0.98 μM IC50 değerleri ile SARS-CoV-2 S aracılı füzyon ve SARS-CoV-2 psödovirüse karşı inhibe edici aktivite gösterdi. Özellikle, EK1, HCoV S’nin HR1 alanını hedefleyen bir pancoronavirüs füzyon inhibitörüdür. SARS-CoV-2 S2 alt birimi HR1 ve HR2 alanlarının 6-HB çekirdeğinin X-ışını kristal yapısı, birkaç mutant kalıntısının olduğunu göstererek çözülmüştür. HR1 bölgesindeki HR2 bölgesindeki gelişmiş etkileşim ile ilgili olabilir. Daha sonra, EK1’den türetilen bir lipopeptit olan EK1C4 üretildi ve SARS-CoV-2 S aracılı hücre-hücre füzyonunu inhibe ettiği doğrulandı. Beklendiği gibi, SARS-CoV-2 S psödovirüs girişi, orijinal EK1 peptidinden ~ 149 kat daha güçlü olan 15,8 nM IC50 ile EK1C4 tarafından da inhibe edildi. Başka bir sekans bazlı lipopeptid füzyon inhibitörü olan IPB02, SARS-CoV-2 S protein aracılı hücre-hücre füzyonunu ve psödovirüs enfeksiyonunu güçlü bir şekilde inhibe eder.
Peptid füzyon inhibitörlerine ek olarak, şu anda reçete edilen bir anti-HIV proteaz inhibitörü olan nelfinavir mesilat (Viracept), hem SARS-CoV-2 S hem de SARS-CoV S aracılı hücre-hücre füzyonunu baskılar. Viracept, peptid füzyon inhibitörlerine ek olarak bildirilen ilk küçük moleküllü füzyon inhibitörüdür. Ayrıca nelfinavir, S proteininin aktivasyonunda rol oynayan TMPRSS2’nin fonksiyonunu inhibe edebilir. Bu keşif, anti-SARS-CoV-2 terapötiklerinin, özellikle enfeksiyonun erken aşamasında, olası klinik uygulamalarını sağlar.
SARS-CoV-2 S bölünme bölgelerini hedefleyen proteaz inhibitörleri
SARS-CoV-2 girişi, S1 / S2 ve S2 bölgelerinde S proteininin bölünmesini gerektirir. TMPRSS2 ve katepsin B ve L ile yapılan proteoliz, SARS-CoV-2 S’nin giriş için hazırlanmasında önemli bir rol oynar. Camostat mesilat, TMPRSS2’nin güçlü bir serin proteaz inhibitörüdür. SARS-CoV ve SARS-CoV-2 hücre giriş mekanizması üzerine yapılan araştırmalardan yararlanılarak, SARS-CoV-2 hücresel girişinin kamostat mesilat tarafından bloke edilebileceği gösterilmiştir. Camostat mesilatın etkinliğini değerlendirmek için şu anda kayıtlı beş klinik çalışma bulunmaktadır (ClinicalTrials.gov Tanımlayıcı: NCT04321096, NCT04353284, NCT04338906, NCT04355052, NCT04374019). Ek olarak, lizozomlardaki katepinler, endositoz yoluyla SARS-CoV girişi için çok önemlidir. Bir katepsin L inhibitörü olan E-64d, SARS-CoV ve SARS-CoV-2 PsV ile enfeksiyonu engeller. COVID-19 hastaları ile gelecekteki denemeler, E-64d tedavisinin etkinliğini doğrulamaya yardımcı olabilir.
Fosfatidilinozitol 3-fosfat 5-kinaz (PIKfyve), erken endozomlarda PI (3,5) P2 sentezleyen ana enzimdir. PIKfyve35’in güçlü bir inhibitörü olan Apilimod, SARS-CoV S psödovirüsünün erken endozomlar aracılığıyla 293 / hACE2 hücrelerine doza bağımlı bir şekilde girişini önemli ölçüde azaltabilir. 293 / hACE2 hücrelerini başka bir PIKfyve inhibitörü olan YM201636 ile tedavi etmek de benzer bir etkiye sahipti. Ayrıca, iki gözenekli kanal alt tipi 2 (TPC2) olan PI (3,5) P2’nin büyük bir aşağı akış efektörü SARS-CoV-2 girişi için önemlidir ve tetrandrin (bir TPC2 inhibitörü) SARS-CoV aktivitesini inhibe eder. -2 S psödovirüs.
Furin (proprotein konvertaz (PC) subtilisin kexin 3, PCSK3), PC ailesinin bir üyesi olarak, çiftli bazik artıklarda peptit ve protein substratlarının hidrolizini katalize eder. Çarpıcı bir şekilde, SARS-CoV-2 S, SARS-CoV-2 iletiminin etkinliğini artırabilecek S1 / S2 sınırında bir furin bölünme bölgesi (682-685 kalıntı) barındırır. SARS-CoV-2’nin S proteinindeki furin benzeri bölünme bölgesi, viral yaşam döngüsü ve patojenite için etkilere sahip olabilir. Bu nedenle furin inhibitörleri, SARS-CoV-2 için bir ilaç tedavisi olarak kullanılabilir. 1994’ten beri patent literatürü, furin veya inhibitörlerinin hastalıkların tedavisinde kullanımını ve a-1-PDX (α1-antitripsin Portland), heksa-D-arginin (D6R), serpin dahil olmak üzere bildirilen bazı furin inhibitörlerini açıklamaktadır. proteinaz inhibitörü 8 (PI8) ve bir peptidomimetik furin inhibitörü.
SARS-Cov-2 S proteinini hedefleyen antivirüs ilaçlarının geliştirilmesinin gelecekteki yönleri
SARS-CoV-2 S proteini, konakçı hücre reseptörüne bağlanır ve virüs istilası sürecinde hayati bir rol oynayan virüs-hücre membran füzyonunu indükler. Ayrıca, S proteini ile ACE2 arasındaki yüksek afinite, SARS-CoV-2’nin bulaşıcılığını arttırır. Pangolinler, evcil hayvanlar (köpekler ve kediler) dahil olmak üzere memeliler, SARS-CoV ve SARS-CoV-2’den S ile ilişki için anahtar kalıntıları belirlemek için önemlidir. SARS-CoV-2 S’nin yapısının ve işlevinin daha iyi anlaşılması, virüsün istilası ve patogenezi hakkında ek bilgi sağlayacaktır. Bu, antiviral terapötiklerin keşfini ve hassas aşı tasarımını destekleyecektir.
Yapısal bilgi, SARS-CoV-2 S proteininin mutasyonlarının değerlendirilmesine de yardımcı olacaktır. Bu kalıntıların, diğer koronavirüslerden S proteinlerinin bilinen antikor epitoplarına yüzey maruziyetine sahip olup olmadığının belirlenmesine yardımcı olacaktır. Ek olarak, yapısal bilgi, yapılar tarafından üretilen proteinlerin homojen olmasını ve insan mAb’lerini nötralize etmek için bir aday aşı veya B-hücresi probu olarak kullanıldığında en nötralizasyona duyarlı epitopları muhafaza etmesi gereken ön füzyon konformasyonuna katılmasını sağlar. Ayrıca atom düzeyinde ayrıntılar, füzyonu engelleyen küçük moleküllerin tasarımını ve taranmasını sağlayacaktır. SARS-CoV-2 ve SARS-CoV RBD alanları,% 75 amino asit sekans özdeşliği paylaştığından, bu Ab’lerden herhangi birinin yeni ortaya çıkan koronavirüsü nötralize edip etmediğini değerlendirmek için gelecekteki çalışmalar gerekli olacaktır. Genel olarak, SARS-CoV-2’nin S proteini ile ACE2 arasındaki etkileşim, SARS-CoV-2 enfeksiyonu mekanizmasının aydınlatılmasına katkıda bulunmak için daha fazla araştırılmalıdır. Benzer şekilde, S proteininin veya reseptör bağlanma bölgesinin yüksek ekspresyonuna odaklanmak da aşıların geliştirilmesi için büyük önem taşımaktadır.
SARS-CoV-2’nin S2 alt birimi, SARS-CoV S2 alanı ile% 88 sekans homolojisi gösterir ve yapısal olarak korunur. Bu nedenle, bu fonksiyonel motifi hedefleyen antikorların geliştirilmesi, bu iki virüsü ve ilgili CoV’leri çapraz bağlayabilir ve nötralize edebilir. Antiviral peptitler, SARS-CoV-2 membran füzyonunu önler ve potansiyel olarak enfeksiyonun önlenmesi ve tedavisi için kullanılabilir. S2 alt biriminin yüksek oranda korunmuş HR1 alanını hedefleyen EK1C4’ün SARS-CoV-2’ye karşı terapötik potansiyele sahip olmasının beklendiğini belirtmek gerekir. Daha da önemlisi EK1C4, tıbbi özelliklerini artıran, dirence karşı yüksek genetik bariyere sahip olan ve ilaca dirençli mutasyonları kolayca indüklemeyen bir burun damlası olarak kullanılabilir. Diğer yandan, peptid füzyon inhibitörleri klinik olarak yaygın olarak kullanılmayabilir ve düşük biyoyararlanıma sahip olabilir. Bu nedenle, oral küçük moleküllü füzyon inhibitörlerinin geliştirilmesi ana yöndür.
Virüs salgınları seyrinde, dış basınca uyum sağlama yeteneği, virüsün yayılmasını etkileyen önemli bir faktördür. Zarf S proteini ile ilgili olarak, RBD’sinin genindeki rekombinasyon veya mutasyon, farklı konakçılar arasında iletimi teşvik etmek ve daha yüksek bir ölüm oranına yol açmak için meydana gelebilir. Aspartatın (D) pozisyon 614’te glisin (G614) mutasyonu, daha patojenik bir SARS-CoV-2 suşu ile sonuçlanır, bu da koruyucu olmayan bölgeleri hedefleyen antikorlar veya aşılar geliştirmeyi daha zor hale getirir. Hastalığı etkili bir şekilde önlemek için, SARS-CoV-2 S yüzeyinde farklı epitopları tanımlayan farklı mAb’lerin kombinasyonları, kaçış mutantı dahil olmak üzere çok çeşitli izolatları nötralize etmek için değerlendirilebilir.
Şu anda, SARS-CoV-2 enfeksiyonunu tedavi etmek veya önlemek için klinik olarak hiçbir spesifik terapötik veya profilaktik kullanılmamıştır. Çin’de COVID-19’u tedavi etmek için IFN-α (rekombinant insan IFN-α1b, IFN-α2a), remdesivir, klorokin, favipiravir ve lopinavir-ritonavir (Aluvia) gibi spesifik olmayan antiviral ilaçlar klinik olarak kullanılmıştır. Bununla birlikte, NIAID-VRC bilim adamları, mRNA aşı platformu teknolojisinde SARS-CoV-2 S proteinini ifade eden aday bir aşı geliştiriyorlar. Aşının klinik denemelerinin önümüzdeki aylarda yapılması bekleniyor. SARS-CoV-2 S proteininin izlenmesinin sürekli olarak güçlendirilmesi, sonraki yeni ilaç geliştirme ve COVID-19’a karşı koruma için büyük önem taşımaktadır.
Şiddetli akut solunum sendromu koronavirüsü 2 (SARS-CoV-2), koronavirüs hastalığına (COVID-19) neden olur. Virüs, pozitif zincirli RNA [(+) RNA] virüslerine aittir. Bunlar Zika, hepatit C (+) RNA virüslerini içeren geniş bir virüs sınıfıdır. (+) RNA virüsleri, genomlarını bulaşıcı viriyonlarında haberci-anlamlı RNA (mRNA) olarak paketler ve bu genomları yalnızca replikasyon komplekslerindeki (RC’ler) RNA ara maddeleri aracılığıyla çoğaltırlar. RC’ler, hücre içi zarların yeniden düzenlenmesiyle oluşturulur. RNA replikasyonu, “remdesivir” dahil antiviral ilaçların ana hedefidir. Koronavirüs ve diğer bazı (+) RNA virüslerinin RC’leri ∼250 ila 300 nm çapında çift zarlı veziküller (kesecikler)dir (DMV’ler). DMV’ler, viral çift sarmallı RNA (dsRNA) replikasyon ara ürünlerini içerir.
Yeni bir çalışma, SARS-CoV-2 ve diğer koronavirüs DMV’lerinde taç benzeri çift membran (zar) enince uzanan moleküler gözenekler (por) tanımladılar. Bu keşif, muhtemelen yeni (+) RNA genomlarının DMV’lerden nasıl salındığı konusunda uzun süredir var olan soruya cevap da oluşturabilir.
Diğer (+) RNA virüsleri gibi, SARS-CoV-2 genomunun çoğu (∼% 70) RNA replikasyon (kendini çoğaltma) fonksiyonlarını kodlar. RC’ler, viral RNA replikasyon proteinlerini, viral RNA şablonları, RNA replikasyonu için gerekli spesifik konak faktörlerini ve ardışık çoğalma aşamalarını düzenleyerek genom replikasyonunu destekler.
Koronavirüs enfeksiyonu çeşitli tipte membran düzenlemelerine yol açsa da, çok sayıda kanıt, dsRNA içeren DMV’leri viral RNA sentez siteleri olarak tanımladı. Bununla birlikte, DMV’ler bilinen açıklıklardan (por) yoksun olduğu için, DMV’nin iç kısmındaki dsRNA’dan kopyalanan yeni (+) RNA genomlarının, viriyonlara paketlenerek ve potansiyel olarak yeni RC’ler oluşturarak sitoplazmaya nasıl geçebildiği açık değildi.
Yeni bir çalışma, SARS-CoV-2 ve başka bir koronavirüs ile enfekte olmuş hücrelerde DMV çift membranlarını bir taraftan diğer tarafa geçen silindirik bir protein kompleksini belirlemek için gelişmiş kriyo-elektron tomografisi (kriyo-ET) kullanarak bu bilmece için zorlayıcı bir çözüm sunmaktadır (şekle bakınız) .
Çalışma, bu gözeneğin büyük viral transmembran protein nsp3’ün (yapısal olmayan protein 3) altı kopyasını içerdiğini gösterdi. Nsp3, RNA replikasyonu için gereklidir ve viral nsp4 ile DMV’lerin oluşumunu indükler. Nsp3’ün çoklu viral replikasyon proteinleri ile etkileşimiyle tutarlı olarak, araştırmacılar, gözeneğin (yani porun) DMV lümen ve sitoplazmik taraflarının diğer makromoleküller ile sıkı ve dinamik etkileşimini görüntülediler. Dolayısıyla, por, yeni sentezlenen RNA’ları sitoplazmaya yönlendirmek için viral RNA polimeraz ve diğer lüminal RNA replikasyon faktörleri ile etkileşime girebilir. Sitoplazmada nsp3’ün viral nükleokapsid proteini ile etkileşimi, RNA’nın yeni viryonlara paketlenmesini kolaylaştırabilir.
Araştırmacılar, koronavirüsün bu por’unun sitozolik kısmını bir “taç” olarak adlandırıyorlar. Bununla birlikte, bu hücre içi DMV RC taçı, koronavirüslere isimlerini veren viryon zarf Spike proteinlerinin taç benzeri yapısı ile ilgisi yoktur.
Replikasyonun karmaşık tacı: RNA replikasyon kompleksleri (RC’ler), çift sarmallı RNA (dsRNA) içeren farklı zar bölmeleridir. Koronavirüs RC’lerinde taç benzeri gözenek (por) kompleksi, yeni yapılmış RNA’ları sitoplazmaya salmak için bir kanal sağlar.
Böylece, yeni genomik RNA’ların taç aracılı salınımı, yaygın bulunan (+) RNA virüsü patojenlerinin farmakolojik veya genetik yollarla kontrolü için bir temel sağlayabilir.
Sitozole açılan bir kapı: Moleküler gözenek (por), koronavirüs replikasyon organelinin çift zarını boru gibi geçer
Koronavirüsler, konakçı hücre zarlarını özel çift zarlı veziküllere (kesecik) dönüştürür. Bu veziküllerin uzun süredir viral genom replikasyonunu barındırdığı düşünülmektedir. Ancak bu kesecikler tamamen kapalı olduğundan, bunların içinde yeni yapılan viral RNA’lar ifade edilme ve yeni viriyonlara paketlenmek üzere sitozole nasıl geçebiliyorlar? Yeni bir kriyo(soğutmalı)-elektron mikroskobu çalışması, çift zarı kapsayan bir moleküler gözenek (por) tanımladı. Bu yapının çekirdeğini büyük bir koronavirüs transmembran proteininin (Nsp3) altı kopyası oluşturuyor. Yapı, bir viral RNA gönderme kanalı olarak kullanılabilir ve böylece gelecekteki antiviral müdahaleler için bir hedef sağlayabilir.
SARS-CoV-2 son 20 yıl içinde insanlarda potansiyel olarak ölümcül bir koronavirüs enfeksiyonunun üçüncü ve en etkili örneğidir (diğerleri SARS ve MERS). Koronavirüsler, pozitif sarmallı RNA (+ RNA) virüsleridir. Alışılmadık derecede büyük genomlarını konakçı hücrenin sitoplazmasında kopyalarlar. Bu süreç, viral replikasyon organeli (RO) olarak bilinen, virüs kaynaklı dönüştürülmüş endoplazmik retikulum (ER) membranları ağı tarafından desteklenir. Çift membranlı veziküller (DMV’ler), RO’nun en bol bileşenidir ve viral RNA sentezi için merkezi öneme sahiptir. DMV’nin içi çift sarmallı (ds) RNA biriktirir. Bunlar muhtemelen viral genom replikasyonunun ve subgenomik mRNA sentezinin ara ürünleridir. DMV’ler, viral RNA sentezi için uygun bir mikro ortam sunabilir ve viral RNA’yı, dsRNA tarafından aktive edilen doğal bağışıklık sensörlerinden koruyabilir. Koronaviral DMV’ler, sitozole açıklıkları olmayan kapalın bölmeler (kompartmanlar) olarak karakterize edilmiştir. Bununla birlikte, yeni yapılan viral mRNA’ların çeviri (proteinlere ifadesi) için dışa (sitoplazmaya) aktarılması gerekir. Ayrıca, koronavirüs genomunun, salgı yolu membranları üzerindeki virüs birleşme bölgelerine hedeflenmeden önce sitozolik nükleokapsid (N) proteini tarafından paketlenmesi gerekir.
Cryo-ET ile ortaya çıkan koronavirüs kaynaklı DMV’ler: (A) Bir kriyo-lamelin tomografik dilimi (7 nm kalınlığında), enfeksiyonun orta aşamasında MHV ile enfekte bir hücreden alınmıştır. (B) Bölümlere ayrılmış içeriğin açıklandığı üç boyutlu (3D) tomogram modeli.
DMV membranlarına gömülü moleküler gözeneklerin mimarisi: Tomografik dilimler (7 nm kalınlığında), gözenek (por) komplekslerinin hem (A) MHV kaynaklı DMV’lerde, hem de (B) SARS-CoV-2 ile indüklenen DMV’lerde (beyaz ok başları) mevcut olduğunu ortaya çıkardı. (A) ‘daki yakınlaştırma, beyaz parantezlerle gösterilen alanın yakından görünümüdür. (C) Merkezi dilim, sivri uçların yakınında (siyah ok başı) ve DMV lümen tarafında esnek veya değişken kütlelerin varlığını düşündürmektedir. (D’den F’ye) Dış (sarı) ve iç (mavi) DMV membranlarına gömülü gözenek kompleksinin (bakır renkli) 3B yüzey oluşturulmuş modelinin farklı görünümleri. (G’den L’ye) Farklı yüksekliklerde gözenek kompleksi boyunca 2D kesit dilimleri. (M ve N) Altı katlı simetrik hacmin (c6, yeşil) altındaki ek bir yoğunluk, simetrik olmayan ortalamada (c1) merkez dışı asimetrik yoğunluk olarak görünüyor.
Koronavirüsler, iki büyük replikaz poliproteini ifade eder. Bu poliproteinler proteolitik olarak 16 yapısal olmayan proteine (nsps) bölünür. Bu nsps’den üçü – nsp3 (MHV’de 222 kDa), nsp4 (56 kDa) ve nsp6 (33 kDa) – transmembran proteinlerdir ve dolayısıyla gözeneğin potansiyel bileşenleridir. Bu nspler, sırasıyla iki, dört ve altı transmembran alanı (TMD’ler) içerir (Şekil 3A) ve çeşitli homotipik ve heterotipik etkileşimlere girer ve çift membranlı RO’lerin oluşumunu yönlendirirler. Nsp3’ün N-terminal kısmının alan bileşimi ve boyutu, koronavirüsler arasında oldukça değişkendir. Birkaç nsp3 alanı (N-terminal ubikuitin benzeri alan 1 ve N proteini) viral RNA ile etkileşime girebilir.
Koronavirüs transmembran proteini nsp3, gözenek kompleksinin bir bileşenidir: (A) (Üst) MHV transmembran nsps’nin membran topolojisi, proteaz bölünme bölgeleri turuncu (PL1pro), kırmızı (PL2pro) ve gri (Mpro) ok uçları ile gösterilir. (Alt) Nsp3’ün, alt alanlarından bazılarını ve MHV-Δ2-GFP3’te bulunan ek EGFP parçasının konumunu gösteren ayrıntılı tasviri. PLpro, papain benzeri proteaz; Mpro, ana proteaz. (B) Gömülü gözenek kompleksleri (beyaz ok başları) ile MHV-Δ2-GFP3 tarafından indüklenen DMV’lerin tomografik dilimi. (C ve D) (C) vahşi tip (wt) MHV ve (D) MHV-Δ2-GFP3 tarafından indüklenen DMV’lerdeki por komplekslerinin altı-katlı simetrik alttomogram ortalamalarının merkezi dilimlerinin karşılaştırılması. (E) Mutant ve yabanıl tip yapılar arasındaki, ikincisi üzerinde yeşil bir kaplama olarak gösterilen 3 standart sapmanın yoğunluk farkları, mutant kompleksinde (siyah ok başları; ayrıca bkz. Film S3) ek (EGFP) kütlelerin varlığını ortaya çıkardı.
Modelde, spesifik replikaz alt birimleri, yeni sentezlenen RNA’yı ona doğru yönlendirmek için gözenek kompleksiyle ilişkilendirilebilir (Şekil 4A). Diğer + RNA viral RO’lar için önerildiği gibi, yalnızca + RNA’ların dışa aktarılması gerekirken, negatif sarmallı şablonlar ve dsRNA ara ürünleri DMV’lerin içinde kalabilir. Sitozolik tarafta, dışa aktarılan tüm viral mRNA’lar N proteini ile birleşebilir (Şekil 4B). Alternatif olarak, biriken N proteini, paketleme için yeni yapılmış genomların bir kısmının seçilmesine hizmet edebilir. Geri kalanı daha sonra çok daha küçük, ancak çok daha bol olan subgenomik mRNA’larla birlikte ifade için kullanılacaktır. Genom içeren RNP kompleksleri, viral zarf proteinlerinin biriktiği ve yeni yapılan viryonlarının bir araya geldiği zarlara gidecektir (Şekil 4C). Bunlar, tipik olarak ER-Golgi ara bölmesinden türetilen tek zarlı bölmelere (Şekil 4D) tomurcuklanır ve hücre dışı boşluğa salınmak üzere salgı yolu boyunca ilerler.
DMV lümeninden (boşluk) virüs tomurcuklanma bölgelerine geçiş yapan koronavirüs genomik RNA modeli: MHV ile enfekte olmuş hücrelerden (üstte) alınan tomografik dilimler, modeldeki (alt) ilgili adımları vurgulamaktadır. (A) Moleküler gözenek (por), viral RNA’yı, N proteini tarafından kapsüllenebileceği sitozole (B) aktarır. (C) Sitozolik RNP kompleksleri daha sonra zar birleşmesi ve (D) viryonların müteakip tomurcuklanması için virüs birleşme bölgelerine gidebilir. Üst panellerdeki girintiler, beyaz parantezlerle gösterilen alanların yakından görünümleridir.
Burada ortaya çıkan çift zarın bir tarafından diğer tarafına yayılan moleküler por, büyük ve çok işlevli nsp3 merkezi bileşeni olmak üzere, DMV’nin iç kısmından sitozole doğru koronaviral RNA ürünleri için çıkış yolunu oluşturabilir. Bu moleküler gözeneğin tam işlevi bilinmese de, viral replikasyon döngüsünde muhtemelen koronavirüsler arasında korunan ve dolayısıyla koronavirüse özgü bir ilaç hedefi sunabilen anahtar bir yapı gibi görünüyor.













 Coronavirusün yeni tacı!
Coronavirusün yeni tacı!



